
发表期刊: PNAS
发表时间: 2019-11
影响因子:9.58
HNF-1β是一种组织特异性转录因子,它与肾脏正常发育和肾小管功能密切相关。HNF-1β的突变会引发许多疾病,比如糖尿病,肾囊肿,癌症等,HNF-1β可直接调控肾囊肿相关基因的表达,控制肾脏发育相关途径。经典Wnt信号通路是由β-catenin介导的Wnt信号途径,由于Wnt配体和细胞表面受体结合,导致β-catenin在细胞核内积累,并与TCF/LEF家族转录因子形成复合体激活Wnt靶基因的转录。目前关于HNF-1β对Wnt信号通路的调控作用尚未有研究,因此,本文利用RNA-seq和ChIP-seq技术,对肾上皮细胞的Wnt靶基因进行分析,从而揭示转录因子HNF-1β通过与β-catenin/LEF竞争性结合染色质,来抑制Wnt靶基因的转录机制。
1. RNA-seq揭示HNF-1β的缺失激活了体内和体外Wnt信号通路
为了研究HNF-1β缺失对Wnt信号通路的影响,及其对依赖于β-catenin基因转录的影响,作者选取了Wnt3a(Wnt配体)处理和不做任何处理的,HNF-1β缺失细胞(KO)、mIMCD3野生型细胞(WT)共4组样本分别进行RNA-seq(图1A)。RNA-seq结果显示,Wnt3a处理后,WT细胞中有124个基因表达发生改变,而KO细胞中表达发生变化的基因数量是WT细胞的8倍(图1A),并且与WT细胞相比,KO细胞中表达发生改变的基因数量和变化幅度均明显增加。另外,Wnt靶基因Axin2是上调幅度最大的基因,通过qRT-PCR验证,Wnt3a处理后,Axin2表达在WT细胞中上调110倍,在HNF-1β KO细胞中显著上调420倍 (图1 B)。这些结果均表明,HNF-1β缺失细胞对Wnt3a蛋白反应强烈。
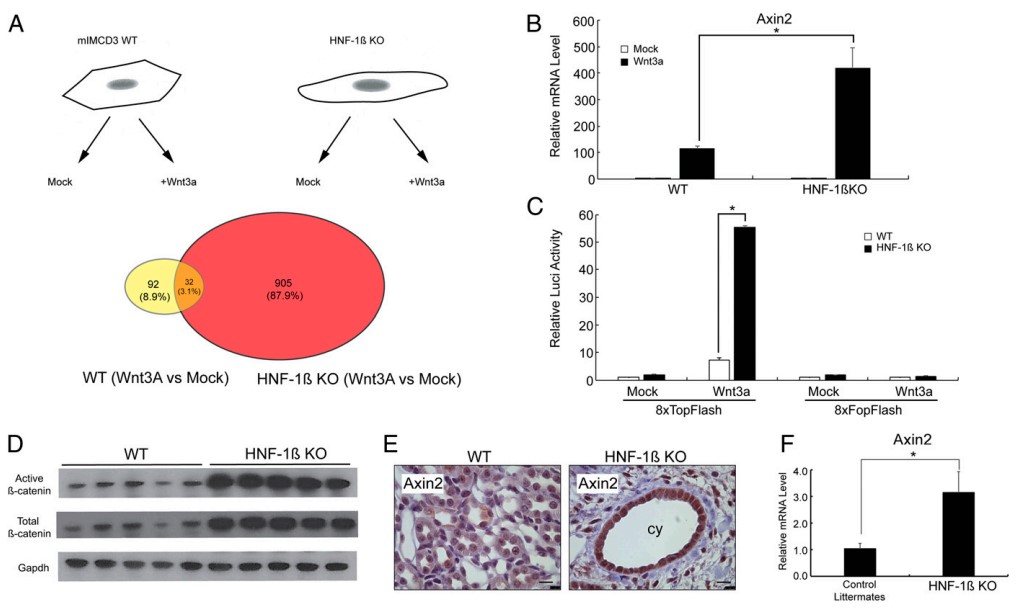
图1 HNF-1β缺失肾上皮细胞和HNF-1β突变肾脏中典型Wnt信号通路的激活
通过荧光素酶报告基因实验,将TOPFLASH(含有TCF/LEF结合位点)和FOPFLASH(TCF/LEF结合位点突变)的报告基因质粒分别转染到野生型细胞(WT)和HNF-1β缺失细胞(KO)中,发现Wnt3a处理后,转染TOPFLASH报告基因质粒的WT细胞荧光素酶活性增加了7倍,KO细胞增加了55倍;而转染FOPFLASH质粒的WT和KO细胞荧光素酶活性均无变化(图1C),这说明HNF-1β缺失细胞对Wnt3a蛋白反应强烈。为了进一步确定HNF-1β突变激活了体内Wnt信号通路,作者又做了以下验证实验。WB结果显示,与同窝出生野生型小鼠比较,出生28天后HNF-1β已敲除的小鼠肾脏溶菌产物中脱磷酸β-catenin蛋白和总β-catenin蛋白含量均增加(图1D)。IHC结果显示,突变肾脏的囊肿上皮细胞中Axin2蛋白明显增加(图1E)。qRT-PCR结果表明,HNF-1β突变肾脏Wnt靶基因Axin2的表达量增加了3倍(图1F)。综上,肾脏上皮细胞HNF-1β的缺失导致体内Wnt信号通路被过度激活。
2. ChIP-seq揭示HNF-1β和β-Catenin与染色质相互结合
为了解释HNF-1β突变导致Wnt信号通路被激活的原因,作者先是选用β-catenin抗体,对Wnt3a处理和不做任何处理的HNF-1β突变肾上皮细胞(KO)、野生型细胞(WT)共4组样本分别进行ChIP-seq(图2A),通过检测β-catenin与染色质结合位点,分析依赖于活性β-catenin的基因转录情况。结果表明,Wnt3a处理后,WT细胞检测到了203个peaks,HNF-1βKO细胞β-catenin的结合峰信号最强,共检测到了1122个peaks(图2A,B)。可见,经Wnt3a处理后,HNF-1β突变细胞β-catenin结合位点最多,这也解释了基因表达被激活的机制。
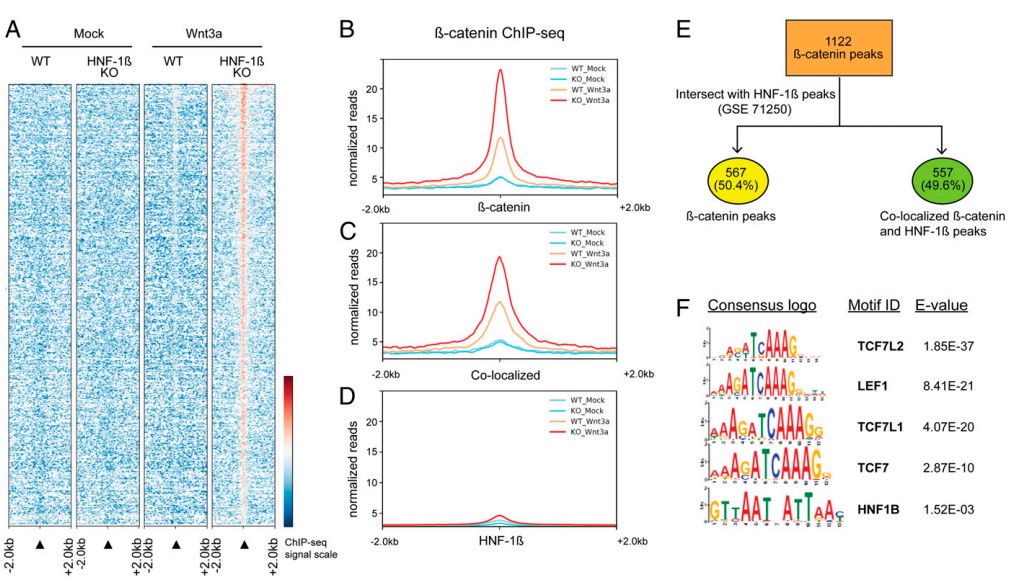
图2 HNF-1β和β-catenin在肾上皮细胞全基因组范围内的共同结合位点接着,作者又选用HNF-1β抗体进行ChIP-seq,检测到28614个HNF-1β结合位点,通过比较HNF-1β突变细胞中β-catenin的结合位点与野生型细胞中HNF-1β的结合位点,发现HNF-1β突变细胞几乎一半的β-catenin结合位点(557/1122peaks;49.6%)与野生型细胞的HNF-1β结合位点相同(图2C,E)。以上结果表明肾上皮细胞的β-catenin和HNF-1β与染色质相互结合。对这557个共同结合位点进行Motif分析,发现TCF/LEF的一些motifs显著富集(图2F),而β-catenin与TCF/LEF在基因组上结合位点又相同;另外也鉴定到HNF-1β和HNF-1α的一些显著富集motifs,说明这557个peaks中存在β-catenin/LEF和HNF-1β的共同结合位点。大多数共同结合位点位于内含子或基因间区,说明它们可能在Wnt信号通路的激活过程中起着顺式调控元件的作用。综上,这些结果表明HNF-1β直接在基因组水平调节Wnt信号通路。
3. HNF-1β抑制Wnt信号通路的靶基因
为了揭示HNF-1β和β-catenin共同结合位点的功能,作者通过分析比较RNA-seq和ChIP-seq数据,确定这些结合位点是否和依赖于Wnt3a靶基因表达的变化有关。图3A表明Wnt3a导致HNF-1β突变细胞937个基因的差异表达,且β-catenin和HNF-1β结合位点位于genebody区上下游100kb以内。HNF-1βKO细胞中由Wnt3a引发的高度上调表达基因与共同结合位点有关,与HNF-1β单独的结合位点不相关。通过分析大多数上调和下调基因,发现β-catenin和HNF-1β共同结合位点主要位于高度上调基因附近,例如Axin2,Ccdc80和Rnf43等,很少位于下调基因附近(图3A)。并且依赖于Wnt3a基因的表达与HNF-1β的单独结合位点无关。以上结果说明,HNF-1β抑制了依赖于Wnt靶基因的表达,HNF-1β的突变又导致了Wnt靶基因的脱抑制,而这些Wnt靶基因表达与共同结合位点有关。
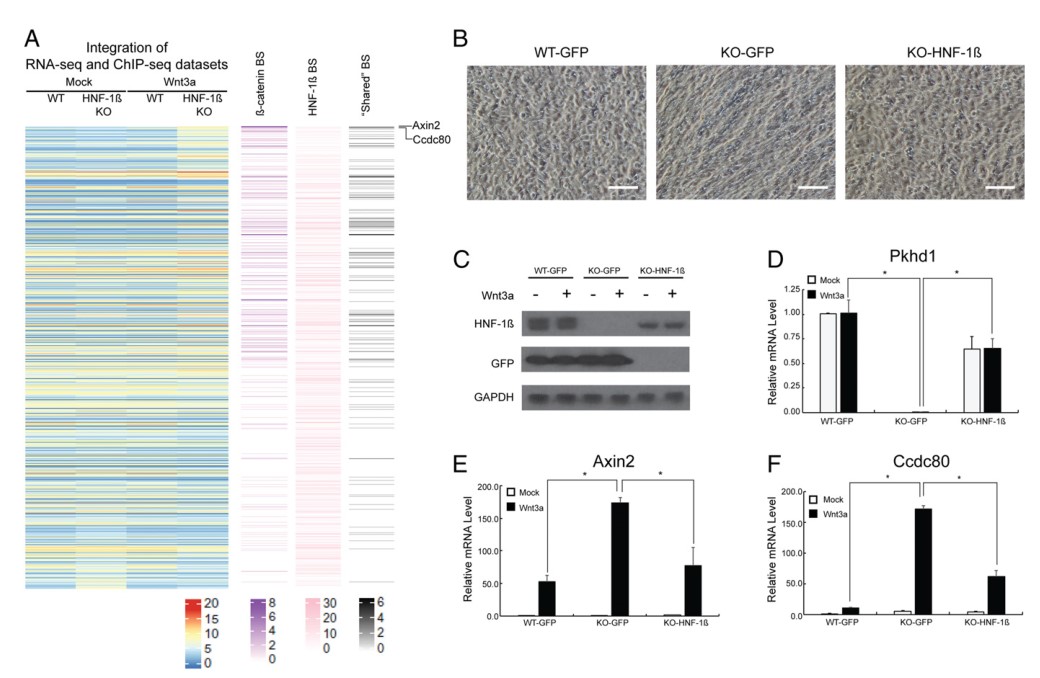
图3 HNF-1β直接抑制典型Wnt信号通路的靶基因
紧接着,作者利用慢病毒让WT细胞的HNF-1β蛋白在HNF-1β突变细胞(KO)中重表达,来验证上述结果。转导GFP后的KO细胞,仍是成纤维细胞样形态;而转导HNF-1β后的KO细胞和转导GFP后的WT细胞相同,均呈现出上皮细胞形态(图3B)。WB结果表明KO细胞HNF-1β蛋白重表达水平同WT细胞相似(图3C)。qRT-PCR结果表明,HNF-1β的消融废除了靶基因Pkhd1的表达,而HNF-1β的重表达修复了Pkhd1的表达,另外Pkhd1表达不受Wnt3a蛋白影响(图3D)。ChIP-seq结果表明,基因Pkhd1 附近存在一些HNF-1β结合位点,而不是单独的或共同的β-catenin结合位点。同野生型细胞相比,Wnt3a处理后,HNF-1β突变细胞中Axin2 和Ccdc80 的表达高度上调(图3E,F),而HNF-1β的重表达减弱了Axin2 和Ccdc80 的转录水平。以上结果表明HNF-1β能够抑制一些典型Wnt信号通路的靶基因表达。
4. β-catenin 和HNF-1β的共同结合峰包含功能增强子靶基因Rnf43,Ccdc80上β-catenin和HNF-1β的共同结合位点分别位于intron2和intron6(图4A-B)。荧光素酶报告基因实验结果显示,Wnt3a处理的HNF-1β突变细胞荧光素酶活性明显高于WT细胞(图4C-F);相反,空载报告基因质粒的荧光素酶活性不受HNF-1β消融或者Wnt3a处理影响。IHC结果显示,HNF-1β突变肾脏的囊肿上皮细胞RNF43和CCDC80蛋白表达均增加,说明靶基因Rnf43,Ccdc80在体内表达上调(图4G-H)。另外,RNA原位杂交结果显示,HNF-1β突变肾脏囊肿上皮细胞中基因Rnf43,Ccdc80的mRNA表达均增加。综上,β-catenin和HNF-1β相互结合,作为顺式调控元件抑制Wnt靶基因的转录,而HNF-1β的缺失又会导致基因转录的脱抑制。
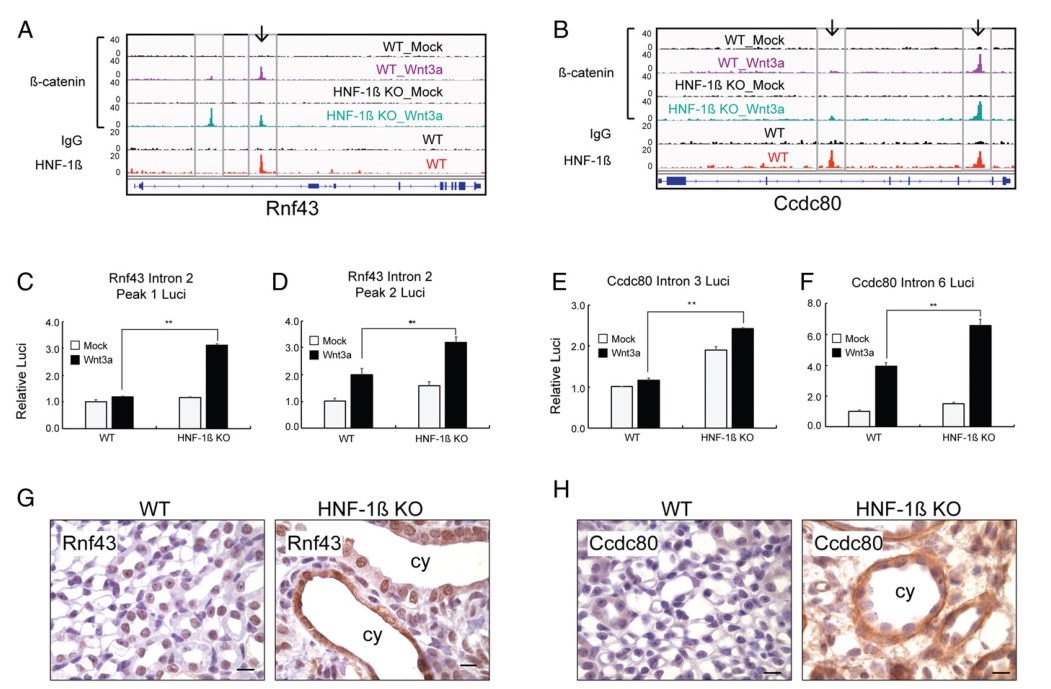
图4 β-catenin 和HNF-1β的共同结合位点包含功能增强子
5. HNF-1β和β-Catenin/LEF竞争结合同一DNA元件
对β-catenin 和HNF-1β的共同结合位点进行深层次的motif分析,发现基因Rnf43 intron2和Ccdc80intron6上β-catenin和HNF-1β的共同结合位点是由一致性的HNF-1β半位点和TCF/LEFmotif重叠组成的复合位点(图5A和D)。
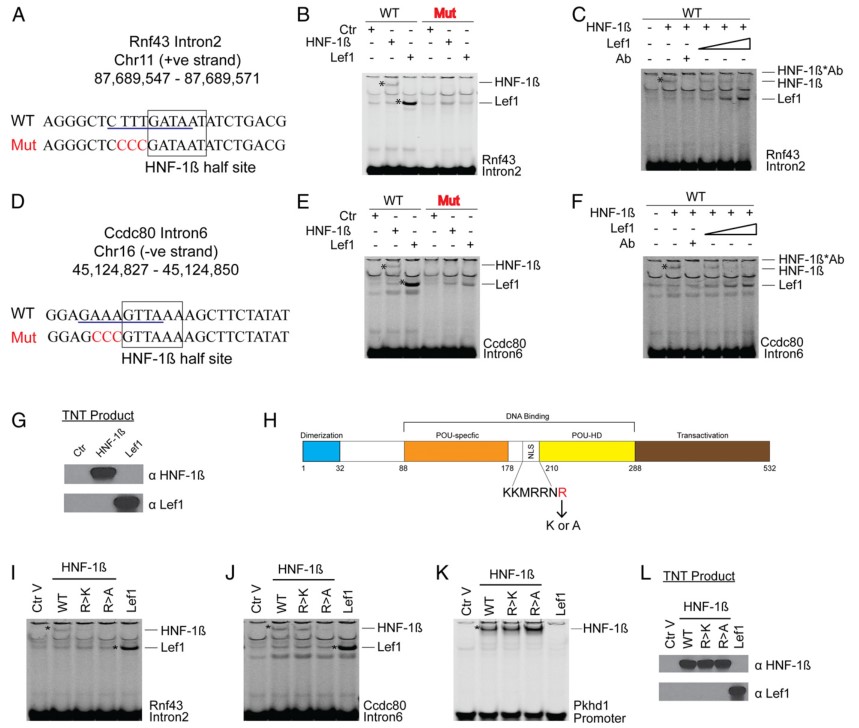
图5 β-catenin 和HNF-1β竞争结合同一复合DNA元件接着,作者借助EMSA实验来验证转录因子TCF/LEF和HNF-1β是否能与基因Rnf43,Ccdc80的复合DNA元件结合,发现LEF1和HNF-1β均能单独结合到复合motif上,而TCF/LEFmotifs的突变导致LEF1和HNF-1β均不能与复合motif结合(图5B和E)。更重要的是,HNF-1β和LEF1在体外能竞争结合复合motif(图5C和F)。另外,HNF-1β半位点的突变导致HNF-1β不能与复合DNA元件结合,也阻止了LEF1在Rnf43位点上的结合,减少了LEF1在Ccdc80位点上的结合,这进一步说明HNF-1β和LEF1竞争结合同一DNA复合元件。由于HNF-1β的DNA结合域C端精氨酸残基突变成赖氨酸或丙氨酸,导致HNF-1β发生突变(图5H)。EMSA结果表明,R209K突变减少了HNF-1β与Rnf43intron2,Ccdc80intron6上复合motif的结合,而R209A突变也废除了HNF-1β的结合(图5I和J)。相反,无论R209R是否发生突变,基因Pkhd1启动子与HNF-1β都有较强结合(图5K和L)。因此,以上结果说明转录因子HNF-1β结合DNA,是通过基因组上HNF-1β和β-catenin共同结合位点的复合motif。
HNF-1β是一种组织特异性转录因子,它对肾脏正常发育和肾小管功能有着重要作用。HNF-1β的突变导致肾囊肿,而肾囊肿与典型Wnt信号通路的反常有关。RNA-seq结果表明,mIMCD3肾上皮细胞中HNF-1β的突变导致其对Wnt3a蛋白反应强烈,同时Wnt靶基因(Axin2、Ccdc80和Rnf43)表达上调。另外,在体内,HNF-1β突变的小鼠肾脏中β-catenin蛋白和Wnt靶基因表达也上调。ChIP-seq结果显示,突变型细胞中HNF-1β的消融导致β-catenin与染色质结合位点数目是野生型细胞HNF-1β的6倍;HNF-1β突变细胞中50%的β-catenin结合位点与野生型细胞HNF-1β结合位点相同,说明HNF-1β与β-catenin在染色质上相互结合。另外,Wnt靶基因Ccdc80和Rnf43均包含由β-catenin/LEF结合位点和HNF-1β半位点重叠组合而成的复合DNA元件,而HNF-1β和β-catenin/LEF共同竞争结合这个元件,因此HNF-1β抑制了依赖于β-catenin基因的转录。综上,本研究揭示了转录因子HNF-1β通过直接抑制β-catenin/LEF与染色质结合,从而限制典型Wnt信号通路的机制。原文链接:https://www.pnas.org/content/116/48/24133

